On May 8, 10:37 am, Jack Linthicum <***@earthlink.net>
wrote:
Phoenician is J2, Aborigines C, figures at the cite
http://www.pnas.org/content/104/21/8726.full
Revealing the prehistoric settlement of Australia by Y chromosome and
mtDNA analysis
1. Georgi Hudjashova,
2. Toomas Kivisilda,b,c,
3. Peter A. Underhilld,
4. Phillip Endicotte,
5. Juan J. Sanchezf,
6. Alice A. Lind,
7. Peidong Sheng,
8. Peter Oefnerh,
9. Colin Renfrewc,i,
10. Richard Villemsa, and
11. Peter Forsterj
+Author Affiliations
1.
aEstonian Biocentre and Tartu University, Department of
Evolutionary Biology, Riia 23, 51010 Tartu, Estonia;
2.
bLeverhulme Centre for Human Evolutionary Studies, University of
Cambridge, Cambridge CB2 1QH, United Kingdom;
3.
dDepartment of Genetics, Stanford University School of Medicine,
Stanford, CA 94305-5120;
4.
eDepartment of Zoology, University of Oxford, Oxford OX1 3PS,
United Kingdom;
5.
fNational Institute of Toxicology and Forensic Science, Canary
Islands Delegation, Campus de Ciencias de la Salud, 38320 La Laguna,
Tenerife, Spain;
6.
gStanford Genome Technology Center, Palo Alto, CA 94304;
7.
hInstitute of Functional Genomics, University of Regensburg,
Josef-Engert-Strasse 9, 93053 Regensburg, Germany;
8.
iMcDonald Institute for Archaeological Research, University of
Cambridge, Downing Street, Cambridge CB2 3ER, United Kingdom; and
9.
jDepartment of Forensic Science and Chemistry, Faculty of
Science and Technology, Anglia Ruskin University, East Road, Cambridge
CB1 1PT, United Kingdom
1.
Contributed by Colin Renfrew, March 30, 2007 (received for
review February 23, 2007)
Next Section
Abstract
Published and new samples of Aboriginal Australians and Melanesians
were analyzed for mtDNA (n = 172) and Y variation (n = 522), and the
resulting profiles were compared with the branches known so far within
the global mtDNA and the Y chromosome tree. (i) All Australian
lineages are confirmed to fall within the mitochondrial founder
branches M and N and the Y chromosomal founders C and F, which are
associated with the exodus of modern humans from Africa ≈50–70,000
years ago. The analysis reveals no evidence for any archaic maternal
or paternal lineages in Australians, despite some suggestively robust
features in the Australian fossil record, thus weakening the argument
for continuity with any earlier Homo erectus populations in Southeast
Asia. (ii) The tree of complete mtDNA sequences shows that Aboriginal
Australians are most closely related to the autochthonous populations
of New Guinea/Melanesia, indicating that prehistoric Australia and New
Guinea were occupied initially by one and the same Palaeolithic
colonization event ≈50,000 years ago, in agreement with current
archaeological evidence. (iii) The deep mtDNA and Y chromosomal
branching patterns between Australia and most other populations around
the Indian Ocean point to a considerable isolation after the initial
arrival. (iv) We detect only minor secondary gene flow into Australia,
and this could have taken place before the land bridge between
Australia and New Guinea was submerged ≈8,000 years ago, thus calling
into question that certain significant developments in later
Australian prehistory (the emergence of a backed-blade lithic
industry, and the linguistic dichotomy) were externally motivated.
* human evolution
* population genetics
Australia was probably occupied by humans at least 50,000 years ago
(1), at a time when lowered sea levels created a land bridge between
Australia and neighboring New Guinea (NG) and when the region was
separated from the Eurasian land mass by only narrow straits such as
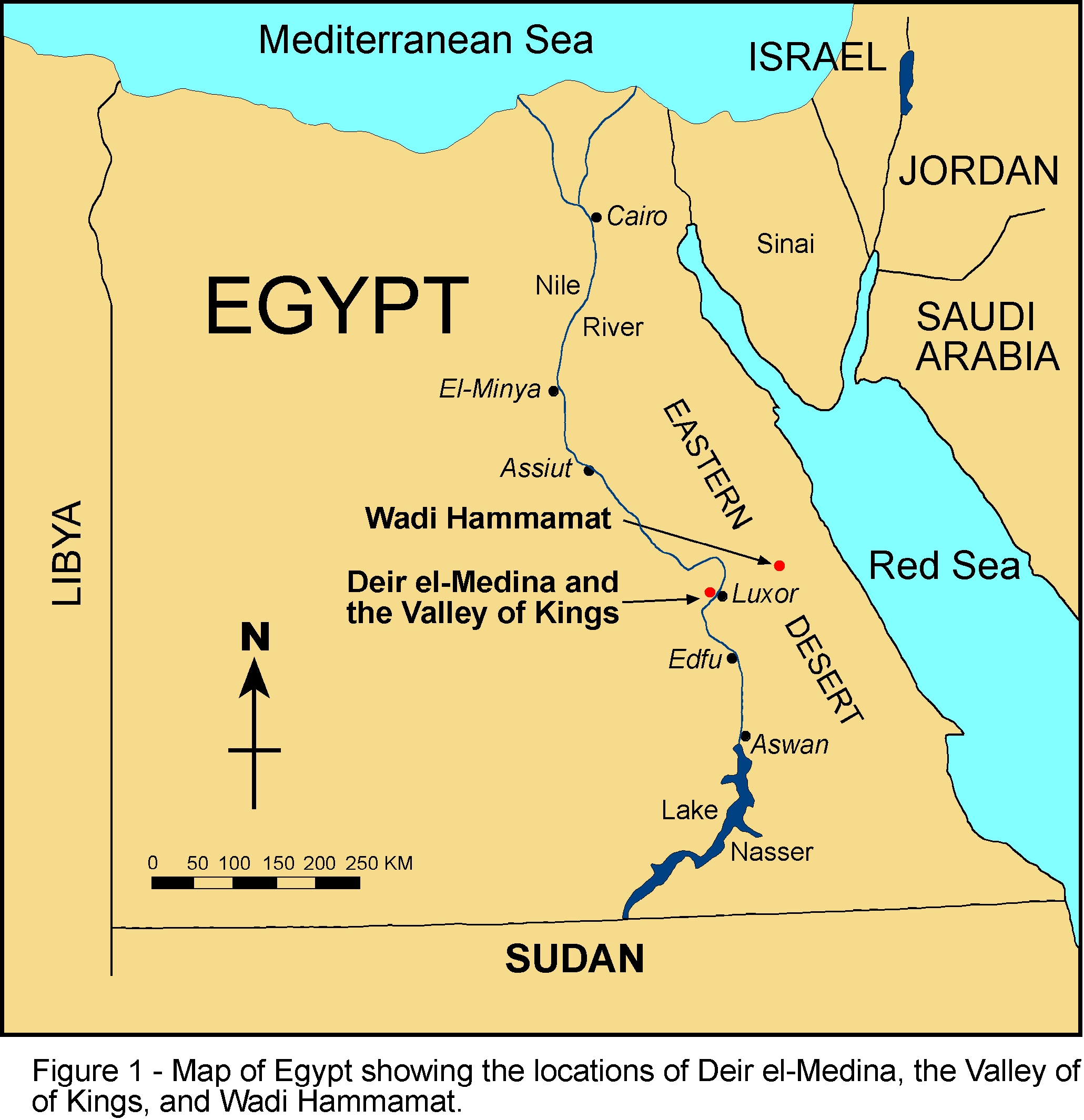
Wallace's Line (Fig. 1). Australia's archaeological record remains
mysterious. To begin with, Australia harbors among the oldest modern
human fossils outside Africa dating to ≈46,000 years (2, 3), despite
the large geographic distance from the African homeland of mankind
(4). Moreover, the earliest known Australian skeletons, at Lake Mungo,
are gracile, whereas some younger skeletal finds (e.g., at Kow Swamp)
have robust morphology (5). Some modern Australian aboriginals retain
elements of this robustness, for example, in the form of pronounced
brow ridges (supraorbital tori) (6, 7). Various explanations can be
put forward for the inconsistent morphological record, for example
that local Homo erectus of Southeast Asia admixed into the modern
human gene pool to a lesser or greater extent (5, 8), or that there
have been multiple migrations to Australia that gave rise to the
differing morphologies at different times, such as hypothetical new
migrants from India (9), or that Australia has been genetically
isolated for a sufficiently long time to produce marked continent-/
Australian-specific features (10–14).
Fig. 1.
View larger version:
* In this page
* In a new window
* Download as PowerPoint Slide
Fig. 1.
Coastlines of Australia and NG ≈50,000 years ago. After the initial
spread of H. sapiens out of Africa to Sahul (the formerly connected
land mass of Australia and NG), the principal processes are
differentiation of the mitochondrial DNA clades Q and S. Subsequent to
that process, there is little migration within Sahul other than Q from
NG to Australia. The genetic isolation of Australia is in the main
very clearly evident already before the Sahul land bridge disappears
≈8,000 years ago. See Results and Discussion for further details.
Archaeological data indicate the intensification of density and
complexity of different stone tools in Australia during the Holocene
period and the emergence of backed-blade stone-tool technology (15).
The first Dingoes (Canis lupus dingo) also appear at about the same
time (3,500–4,000 years ago) and were proposed to have been introduced
by new human arrivals, from India (16), along with new stone tool
types (17). This debate is ongoing (15, 18, 19).
Recent molecular studies on humans have likewise yielded a diversity
of interpretations, ranging from a deep but undated split
distinguishing Australians even from their immediate neighbors to the
north in NG (20) to a very recent immigration event within the
Holocene in the past 10,000 years (21, 22).
Using new Australian and NG samples screened for mtDNA and Y
chromosome variation and benefiting from the increasing genetic sample
coverage available for Australia [Fig. 1 (4, 20–32)], we can now
attempt to clarify some of the salient features of the record of
Australian population history and confirm its considerable isolation.
Previous SectionNext Section
Results and Discussion
African Ancestry of Australian and NG Y and mtDNA Types.
We carried out a phylogenetic analysis of our Australian and NG
complete mtDNA sequences and compared the resulting branches with the
Asian mtDNA tree, as known so far (Fig. 2). The result confirms that
both Australian and NG maternal lineages consist exclusively of the
known out-of-Africa founder types M and N, dated to ≈50–70,000 years
ago, and their derivatives (24–27, 29, 30, 33–37). This mitochondrial
finding is mirrored in our Y chromosome data (Fig. 3), where we
observe the paternal lineages in Australians and New Guineans to fall
into either branches C or F, proposed to be the earliest out-of-Africa
founder types (31). These results indicate that Australians and New
Guineans are ultimately descended from the same African emigrant group
50–70,000 years ago, as all other Eurasians. In other words, these
data provide further evidence that local H. erectus or archaic Homo
sapiens populations did not contribute to the modern aboriginal
Australian gene pool, nor did Australians and New Guineans derive from
a hypothetical second migration out of Africa (38), nor is there any
suggestion of a specific relationship with India (9, 21, 22).
Fig. 2.
View larger version:
* In this page
* In a new window
* Download as PowerPoint Slide
Fig. 2.
Simplified tree of autochthonous Near Oceanian mtDNA branches. East
and Southeast Asian, and Indian specific clusters are added for
comparison. Mutations relevant to Australia, Melanesia, and NG are
shown along the branches. Only branches identified by at least two
complete mtDNA sequences are included. For data and a detailed tree,
see SI Fig. 4.
Fig. 3.
View larger version:
* In this page
* In a new window
* Download as PowerPoint Slide
Fig. 3.
Simplified Y chromosomal phylogeny including the recently discovered
Australia-specific marker M347. For data and a detailed tree, see SI
Fig. 5.
Comparing the Australian complete mtDNA sequences within the context
of the Asian phylogeny (25, 26, 38–45), we find that the Australians
do not share any derived branches with Asians more recent than the
founding types M, N, and R (Fig. 2). Similarly, our increased
resolution of regionally differentiated Y chromosomal types, C5 in
India, C4 in Australia, and C2 in NG provide evidence of significant
long-term isolation (Fig. 3). Although the confirmed existence of F*
chromosomes in India (13, 46) suggests they may also exist in
Australia and NG, incomplete molecular analysis for types G–J in some
previous studies (28, 47) leaves the issue of the presence of basal F*
chromosomes in Australia and NG unresolved. The implication is that
the migration rate of the founders from Africa along the Indian Ocean
has been rapid relative to the mutation rate of the complete mtDNA
genome [one mutation in ≈5,000 years; see Mishmar et al. (37)]. These
findings support the relatively rapid migration of the Eurasian
founder types to Southeast Asia (45) and, as we can now confirm, all
the way to Australia. It should be noted that migration in this
context refers not simply to travel but also to successful
colonization. Applying the given mutation rate to the M, N, and R
founders, the migration from southwestern Asia to Australia would have
taken <5,200 years at 95% confidence, assuming a Poisson mutation
process. This migration speed is in the same order of magnitude as
estimated for other prehistoric continental settlements (48).
Australian and NG Founder Lineages.
An important result in our high-resolution mtDNA data is the discovery
that Australians and New Guineans not only share the same M and N
founders dating from the African exodus but furthermore within M share
a characteristic variant nucleotide position 13500, which is
widespread in Australia, NG, and neighboring Melanesia but not found
elsewhere in the world. Taken together with the fact that the
ancestral node, but not the derived lineages, is shared between
Australia and NG/Melanesia (Fig. 2), we argue for a single founder
group settling the whole region of Australia and NG ≈50,000 years ago.
Strongly supporting evidence for this view comes from the N portion of
the mtDNA phylogeny (Fig. 2), where a major deep subclade P is found
in both Australia and NG/Melanesia but not elsewhere, with the time-
depth estimates for P again ranging around the 50,000-year mark (Table
1).
View this table:
* In this window
* In a new window
Table 1.
Age estimates for mtDNA branches found in Australians, New Guineans,
and Melanesians
Within Australia, the ancient mtDNA branch S (27) stands out, because
it is found in 34% of our Australians [supporting information (SI)
Table 2], and it is well represented in other regions of Australia, as
detailed in Fig. 1 (4, 21, 23–27, 29) and has so far not been detected
elsewhere in the world, based on the available global mtDNA database
of >40,000 sequences (49). This branch is distinguished from the root
of macrohaplogroup N by a transition at nucleotide position 8404.
Nearly all Australians who do not have mtDNA type S nevertheless
harbor deep mtDNA branches specific to Australia, several of which are
described in this study (SI Fig. 4). These deep and continent-specific
branches indicate substantial isolation since the first colonization
of Australia. Although NG and Australia were not separated until 8,000
years ago, we can estimate the time depth for the arrival in Australia
both qualitatively and quantitatively. Qualitatively, as argued above,
the relatively nested phylogenetic structure, with no mutation events
separating the M, N, and R founders around the Indian Ocean even at
the highly resolved level of the complete mtDNA sequence, indicates an
arrival in Australia soon after the African exodus, the latter dated
to 50–70,000 years ago (14, 36, 48). Quantitatively, the absolute date
estimates for the founder clusters in Near Oceania yield dates of up
to 58,000 ± 8,000 years ago (Table 1).
Occurrence of a “New Guinean” Lineage in Northern Australia.
There is an important exception to the general pattern of Australian-
specific lineages in Australian aboriginals, and this concerns mtDNA
branch Q. Thus far, Q has been considered as having a geographic
distribution restricted to NG and Melanesia (25, 26, 30, 33–35, 50).
Surprisingly, in our northern Australian Kalumburu sample, we now find
an Aboriginal Australian mtDNA lineage bearing all of the basic
mutations characteristic of haplogroup Q. This Australian Q lineage
does not appear to be a recent arrival from NG (nor indeed a case of
sample confusion), because the lineage does not belong to any of the
common and widespread Q subclusters known so far from NG and
Melanesia. The Australian Q instead branches deeply within Q to a
depth of five mtDNA mutations. The mutational time of separation of
this Q lineage from existing NG Q branches is estimated at 30,400 ±
9,300 years (Table 1). The geographically restricted appearance of Q
in northern Australia may suggest a secondary arrival of settlers from
NG well before the land bridge between Australia and NG was submerged
≈8,000 years ago.
Apart from this potential signal of secondary migration into
Australia, there seem to be no further lineages either on the
Australian Y or mtDNA tree that would provide clear evidence for
extensive genetic contact since the first settlement, except possibly
for a P3 sublineage shared between Australia and NG (Fig. 2). Thus,
Australia appears to have been largely isolated since initial
settlement, in agreement with one interpretation of the fossil record
(10, 11). In particular, there are no lineages exclusively shared
between Australia and India that might have indicated common ancestry
as originally proposed by Huxley (9). Indeed, we have identified a new
Y marker M347 (Fig. 3), which distinguishes all Australian C types
from Indian or other Asian C types and adds weight to the rejection of
the Huxley hypothesis. NG, in contrast, does carry a clear imprint of
new arrivals at least along its coasts, where the “Austronesian” B
mtDNA type has been established (51, 52).
This conclusion may have a negative bearing on the much-discussed
emergence of a new stone tool industry in Australia, the “small tool”
tradition, characterized by backed blades (15, 53). There is currently
no evidence in Australia to associate this change in the material
culture record with the arrival of new maternal and paternal lineages.
A major question that has not been addressed here and awaits
resolution is the intriguing linguistic landscape of Australia, where
seven-eighths of the continent is dominated by a single language
family (Pama-Nyungan), whereas all other language families are
concentrated in the northwestern region of Australia [Fig. 1 (54)].
Our samples from Kalumburu are from the linguistically diverse
northern zone, where we have identified potential secondary gene flow
into Australia as evidenced by a mitochondrial Q lineage distantly
related to current NG Q lineages. The secondary migration ≈30,000
years ago associated with the arrival of the Q lineage would be
considered too early, in the view of most linguists, to account for
this dichotomy. Future more exhaustive genetic surveys of the
Australian continent may one day resolve whether the Australian
linguistic landscape can be better understood with the identification
of such potential contact events. At present, it may seem preferable
to seek an explanation for the dichotomy in terms of events and
processes internal to Australia.
Previous SectionNext Section
Conclusions
The mitochondrial and Y chromosomal results presented here point
toward one early founder group settling both Australia and NG soon
after the exodus from Africa ≈50–70,000 years ago, at a time when the
lowered sea levels joined the two islands into one land mass,
necessitating sea travel only across narrow straits such as Wallace's
Line. The deep and specific phylogenetic lineages today within this
former landmass indicate a small founding population size and
subsequent isolation of Australia and, to a lesser extent, of NG, from
the rest of the world. These founder events and the lack of contact
could underlie the divergent morphological development seen in the
Australian human fossil record and could also help explain the
remarkably restricted range of Pleistocene Australian lithic
industries and bone artifacts compared with contemporaneous cultures
elsewhere in the world (55).
Previous SectionNext Section
Materials and Methods
Samples.
In total, 172 Australian and Melanesian mtDNAs and 522 Y chromosome
profiles were used in this study. Samples were obtained with informed
consent. The following mtDNA sequences were generated: 32 sampled
Aboriginal Australians from Kalumburu in northwestern Australia and 48
NG highlanders from the Bundi area (Fig. 1). Four of the Australian
individuals had been characterized by Y chromosome short tandem repeat
analysis (20). In addition, mtDNA sequences were generated from the
following DNA samples described in Kivisild et al. (27): two
Aboriginal Australian samples (Oc06 and Oc10), two NG samples (Oc01
and Oc16), and two Melanesian samples (Oc03 and Oc04). Extended Y
chromosomal profiles were generated for the males within these samples
(6/32 Aboriginal Australians and 19/48 New Guineans).
Previously published mtDNA profiles were included as follows: 33
complete or nearly complete mtDNA sequences from Australia (25–27, 29,
30); 32 complete or nearly complete mtDNA sequences from NG (25–27,
30, 33, 34); and 27 complete or nearly complete mtDNA sequences from
Melanesia (25, 27, 30, 34, 35).
Published Eurasian and Near Oceanian Y chromosomal haplotypes in the
present study include: 102 Aboriginal Australians (28, 31, 32); 395
New Guineans (28, 31, 32, 47); 1,021 individuals from Southeast Asian
populations (28, 31, 32, 47, 56); 1,141 individuals from the Indian
subcontinent and Pakistan (13, 31, 32, 46); 358 individuals from East
Asian populations (13, 28, 31, 32); and 1,065 individuals from
Northeast and Central Asian populations (31, 32, 56) (see SI Fig. 5
for further details).
mtDNA Typing.
The first hypervariable segment (HVS-1) of mtDNA (nps 16024–16383) and
the stretch 57–302 of HVS-2 were sequenced directly from both strands
in all samples. Additionally, two macrohaplogroup M and N defining
mutations, namely 10398 A>G and 10400 C>T, were genotyped in all
samples by RFLP (DdeI 10394 and AluI 10397 respectively). M types were
further analyzed, by direct sequencing, for the Q and M29 marker
13500C (33, 50) and N types were checked for the S marker 8404C (27)
and the P marker 15607G (33, 50). Additional coding region markers
were analyzed in S and P mtDNA types (SI Table 3). The haplotypes
defined by control region sequences and coding region SNPs were
further grouped by their mutational motifs under following
subhaplogroups: B4a1a1, M7b1, P1, P2, P3, P4b, Q1, and Q2 (see SI
Table 3 and SI Fig. 4 for further details) (30, 33, 43, 51).
Most NG (40/48) and approximately one-half of Aboriginal Australian
(14/32) mtDNA haplotypes could be sufficiently well characterized
using existing mtDNA haplogroup nomenclature (SI Tables 2 and 3). Of
the 26 mtDNA control region sequences that did not show clear
affiliation to previously described haplogroups, nine Australian and
NG individuals were selected for complete mtDNA sequencing. All
recently characterized mutations that were found during the complete
mitochondrial genome sequencing were typed in individuals with similar
or identical mtDNA control region sequences (SI Table 3).
Multiplex SNP Assay.
A mtDNA multiplex PCR was designed and performed in a reaction volume
of 25 μl containing 1× PCR buffer, 6.5 mM MgCl2, 600 mM each dNTP,
0.01–0.2 mM of each primer (SI Table 4), and 2 units of AmpliTaq Gold
DNA polymerase (Applied Biosystems, Tartu, Estonia). The thermal
cycling program was: denaturation at 95°C for 10 min followed by 35
cycles of 95°C for 30 s, 60°C for 30 s, and 65°C for 30 s, followed by
6 min at 65°C.
Excess primers and dNTPs were removed by addition of 1 μl (1 unit/μl)
of shrimp alkaline phosphatase and 0.02 μl (10 units/μl) of
Exonuclease I (Amersham Pharmacia Biotech, Piscataway, NJ) to 2.5 μl
of PCR product and incubating the mixture at 37°C for 30 min followed
by 80°C for 15 min.
Single-base extension (SBE) reactions were performed in 5 μl with 1 μl
of purified PCR product, 3 μl of SNaPshot (Applied Biosystems), or
SNuPe (Amersham Biosciences, Piscataway, NJ) reaction mix, 0.5 μl of
SBE primer mix (0.01–0.3 mM each primer; see SI Table 5), and 0.5 μl
of water. The SBE primer mix was diluted in 160 mM ammonium sulfate
(Sigma–Aldrich, Helsinki, Finland) to minimize primer-dimer artifacts.
Excess nucleotides were removed by addition of 1 μl (1 unit/μl) shrimp
alkaline phosphatase to the SBE mix and incubation at 37°C for 20 min
followed by incubation at 80°C for 15 min. Two microliters of SBE
product were mixed with 18 μl of Hi-Di formamide (Applied Biosystems)
and 0.1 μl of GeneScan-120 Liz internal size standard (Applied
Biosystems), and analyzed by capillary electrophoresis using ABI Prism
3730XL Genetic Analysers with 50 cm capillary arrays and POP-6 polymer
(Applied Biosystems) or a MegaBACE Analysis System (Amersham
Biosciences). Full methodological and theoretical details are
available elsewhere (57, 58).
Y Chromosome Typing.
Eighteen Y chromosomal markers (M4, M9, M11, M38, M45, M70, M89, M130,
M147, M175, M177, M208, M210, M214, M230, M231, M347, and M356) (SI
Fig. 5) were typed in 25 of the Kalumburu and Bundi samples. One
previously unpublished biallelic M347 marker is reported here. M347
was amplified by using primers (F, 5′-AAGTGGAGGGTATGTTTCAGCC-3′; R, 5′-
GGCAACAATAGGCAGATGGCTC-3′) specific for a single 558-bp amplicon. The
thermal cycling program was: denaturation at 95°C for 3 min followed
by 36 cycles of 95°C for 30 s, 53°C for 30 s, and 72°C for 40 s,
followed by 5 min at 72°C. Nucleotide position 374 A>G
(ancestral>derived) variant was sequenced directly by using one of the
same external primers. The following were additionally genotyped:
haplogroup C* lineages (altogether 13 individuals) from the study by
Kivisild et al. (46) were typed for the presence of the M356 marker
reported by Sengupta et al. (13); NG haplogroup K* lineages from the
study by Underhill et al. (31, 32) were typed for the presence of M230
marker; and the M347 marker was typed in Australian C lineages from
the study by Underhill et al. (31, 32).
Coalescence Age Estimation.
Phylogenetic trees were constructed manually and confirmed by using
the Network software (www.fluxus-engineering.com) (59, 60).
Coalescence ages of mtDNA haplogroups were calculated by the rho (ρ)
statistic as described by refs. 61 and 62 by using the coding region
mutation rate of one synonymous transition per 6,764 years (27).
Previous SectionNext Section
Footnotes
* cTo whom correspondence may be addressed. E-mail:
***@cam.ac.uk or ***@mcdonald.cam.ac.jk
*
Author contributions: T.K., P.A.U., R.V., and P.F. designed
research; G.H., A.A.L., and P.S. performed research; P.E., J.J.S.,
P.O., and P.F. contributed new reagents/analytic tools; G.H., T.K.,
and P.A.U. analyzed data; and G.H., T.K., P.A.U., P.E., J.J.S., C.R.,
R.V., and P.F. wrote the paper.
*
The authors declare no conflict of interest.
*
Data deposition: The sequences reported in this paper have been
deposited in the GenBank database [accession nos. EF495214–EF495222
(complete mtDNA sequences), EF524341–EF524420 (mtDNA HVS-1 sequences),
and EF524421–EF524500 (partial HVS-2 sequences)].
*
This article contains supporting information online at
www.pnas.org/cgi/content/full/0702928104/DC1.
* Abbreviations:
NG,
New Guinea;
SBE,
single-base extension.
* © 2007 by The National Academy of Sciences of the USA
Previous Section
References
1. ↵
1. Stringer C
(2002) Philos Trans R Soc London B 357:563–579.
CrossRefISI
2. ↵
1. Bowler J,
2. Johnston H,
3. Olley J,
4. Prescott J,
5. Roberts R,
6. Shawcross W,
7. Spooner N
(2003) Nature 421:837–840.
CrossRef
3. ↵
1. O'Connell JF,
2. Allen J
(2004) J Arch Sci 31:835–853.
CrossRefISI
4. ↵
1. Vigilant L,
2. Stoneking M,
3. Harpending H,
4. Hawkes K,
5. Wilson AC
(1991) Science 253:1503–1507.
Abstract/FREE Full Text
5. ↵
1. Bräuer G,
2. Mellars P,
3. Stringer C
(1989) The Human Revolution: Behavioural and Biological
Perspectives on the Origins of Modern Humans (Princeton Univ Press,
Princeton, NJ), pp 123–154.
6. ↵
1. Larnach SL,
2. Macintosh NWG
(1970) The Craniology of the Aborigines of Queensland
(University of Sydney, Sydney, Australia).
7. ↵
1. Larnach SL
(1978) Australian Aboriginal Craniology (University of Sydney,
Sydney, Australia).
8. ↵
1. Thorne AG,
2. Wolpoff MH
(1992) Sci Am 266:76–79, 82–3.
9. ↵
1. Huxley TH
(1870) J Ethnol Soc London 2:404–412.
10. ↵
1. Pardoe C
(1991) Arch Oceania 26:79–85.
11. ↵
1. Brown P
(1992) Philos Trans R Soc London B 337:235–242.
CrossRefISI
12. ↵
1. Lahr M
(1996) The Evolution of Modern Human Diversity (Cambridge Univ
Press, Cambridge, UK).
13. ↵
1. Sengupta S,
2. Zhivotovsky LA,
3. King R,
4. Mehdi SQ,
5. Edmonds CA,
6. Chow CE,
7. Lin AA,
8. Mitra M,
9. Sil SK,
10. Ramesh A,
11. et al.
(2006) Am J Hum Genet 78:202–221.
CrossRefMedlineISI
14. ↵
1. Richards MB,
2. Bandelt H-J,
3. Kivisild T,
4. Oppenheimer S
1. Bandelt H-J,
2. Macaulay V,
3. Richards M
(2006) in Human Mitochondrial DNA, the Evolution of Homo
sapiens, eds Bandelt H-J, Macaulay V, Richards M (Springer, Berlin),
pp 225–265.
15. ↵
1. Mulvaney DJ,
2. Kamminga J
(1999) Prehistory of Australia (Smithsonian Institution Press,
Washington, DC).
16. ↵
1. Gollan K
1. Misra VN,
2. Bellwoood P
(1985) in Recent Advances in Indo-Pacific Prehistory, eds Misra
VN, Bellwoood P (Oxford & IBH, New Delhi, India), pp 439–443.
17. ↵
1. Glover IC,
2. Presland G
1. Misra VN,
2. Bellwoood P
(1985) in Recent Advances in Indo-Pacific Prehistory, eds Misra
VN, Bellwoood P (Oxford & IBH, New Delhi, India), pp 185–195.
18. ↵
1. Corbett L
(1995) The Dingo in Australia and Asia (Comstock/Cornell,
Ithaca, NY).
19. ↵
1. Savolainen P,
2. Leitner T,
3. Wilton AN,
4. Matisoo-Smith E,
5. Lundeberg J
(2004) Proc Natl Acad Sci USA 101:12387–12390.
Abstract/FREE Full Text
20. ↵
1. Forster P,
2. Kayser M,
3. Meyer E,
4. Roewer L,
5. Pfeiffer H,
6. Benkmann H,
7. Brinkmann B
(1998) Mol Biol Evol 15:1108–1114.
Abstract
21. ↵
1. Redd AJ,
2. Stoneking M
(1999) Am J Hum Genet 65:808–828.
CrossRefMedlineISI
22. ↵
1. Redd A,
2. Roberts-Thomson J,
3. Karafet T,
4. Bamshad M,
5. Jorde L,
6. Naidu J,
7. Walsh B,
8. Hammer M
(2002) Curr Biol 12:673–677.
CrossRefMedlineISI
23. ↵
1. van Holst Pellekaan S,
2. Frommer M,
3. Sved J,
4. Boettcher B
(1998) Am J Hum Genet 62:435–449.
CrossRefMedlineISI
24. ↵
1. Huoponen K,
2. Schurr TG,
3. Chen Y,
4. Wallace DC
(2001) Hum Immunol 62:954–969.
CrossRefMedlineISI
25. ↵
1. Ingman M,
2. Gyllensten U
(2003) Genome Res 13:1600–1606.
Abstract/FREE Full Text
26. ↵
1. Ingman M,
2. Kaessmann H,
3. Pääbo S,
4. Gyllensten U
(2000) Nature 408:708–713.
CrossRefMedline
27. ↵
1. Kivisild T,
2. Shen P,
3. Wall DP,
4. Do B,
5. Sung R,
6. Davis K,
7. Passarino G,
8. Underhill PA,
9. Scharfe C,
10. Torroni A,
11. et al.
(2006) Genetics 172:373–387.
Abstract/FREE Full Text
28. ↵
1. Kayser M,
2. Brauer S,
3. Cordaux R,
4. Casto A,
5. Lao O,
6. Zhivotovsky LA,
7. Moyse-Faurie C,
8. Rutledge RB,
9. Schiefenhoevel W,
10. Gil D,
11. et al.
(2006) Mol Biol Evol 23:2234–2244.
Abstract/FREE Full Text
29. ↵
1. van Holst Pellekaan SM,
2. Ingman M,
3. Roberts-Thomson J,
4. Harding RM
(2006) Am J Phys Anthropol 131:282–294.
CrossRefMedlineISI
30. ↵
1. Friedlaender JS,
2. Friedlaender FR,
3. Hodgson JA,
4. Stoltz M,
5. Koki G,
6. Horvat G,
7. Zhadanov S,
8. Schurr TG,
9. Merriwether DA
(2007) PLoS ONE 2:e248.
CrossRef
31. ↵
1. Underhill PA,
2. Passarino G,
3. Lin AA,
4. Shen P,
5. Mirazon Lahr M,
6. Foley R,
7. Oefner PJ,
8. Cavalli-Sforza LL
(2001) Annu Hum Genet 65:43–62.
32. ↵
1. Underhill PA,
2. Shen P,
3. Lin AA,
4. Jin L,
5. Passarino G,
6. Yang WH,
7. Kauffman E,
8. Bonné-Tamir B,
9. Bertranpetit J,
10. Francalacci P,
11. et al.
(2000) Nat Genet 26:358–361.
CrossRefMedlineISI
33. ↵
1. Friedlaender J,
2. Schurr T,
3. Gentz F,
4. Koki G,
5. Friedlaender F,
6. Horvat G,
7. Babb P,
8. Cerchio S,
9. Kaestle F,
10. Schanfield M,
11. et al.
(2005) Mol Biol Evol 22:1506–1517.
Abstract/FREE Full Text
34. ↵
1. Pierson MJ,
2. Martinez-Arias R,
3. Holland BR,
4. Gemmell NJ,
5. Hurles ME,
6. Penny D
(2006) Mol Biol Evol 23:1966–1975.
Abstract/FREE Full Text
35. ↵
1. Merriwether DA,
2. Hodgson JA,
3. Friedlaender FR,
4. Allaby R,
5. Cerchio S,
6. Koki G,
7. Friedlaender JS
(2005) Proc Natl Acad Sci USA 102:13034–13039.
Abstract/FREE Full Text
36. ↵
1. Watson E,
2. Forster P,
3. Richards M,
4. Bandelt HJ
(1997) Am J Hum Genet 61:691–704.
MedlineISI
37. ↵
1. Mishmar D,
2. Ruiz-Pesini E,
3. Golik P,
4. Macaulay V,
5. Clark AG,
6. Hosseini S,
7. Brandon M,
8. Easley K,
9. Chen E,
10. Brown MD,
11. et al.
(2003) Proc Natl Acad Sci USA 100:171–176.
Abstract/FREE Full Text
38. ↵
1. Thangaraj K,
2. Chaubey G,
3. Kivisild T,
4. Reddy AG,
5. Singh VK,
6. Rasalkar AA,
7. Singh L
(2005) Science 308:996.
Abstract/FREE Full Text
39. ↵
1. Kong Q-P,
2. Yao Y-G,
3. Sun C,
4. Bandelt H-J,
5. Zhu C-L,
6. Zhang Y-P
(2003) Am J Hum Genet 73:671–676.
CrossRefMedlineISI
40. ↵
1. Palanichamy M,
2. Sun C,
3. Agrawal S,
4. Bandelt H-J,
5. Kong Q-P,
6. Khan F,
7. Wang C-Y,
8. Chaudhuri T,
9. Palla V,
10. Zhang Y-P
(2004) Am J Hum Genet 75:966–978.
CrossRefMedlineISI
41. ↵
1. Kong QP,
2. Bandelt HJ,
3. Sun C,
4. Yao YG,
5. Salas A,
6. Achilli A,
7. Wang CY,
8. Zhong L,
9. Zhu CL,
10. Wu SF,
11. et al.
(2006) Hum Mol Genet 15:2076–2086.
Abstract/FREE Full Text
42. ↵
1. Sun C,
2. Kong QP,
3. Palanichamy MG,
4. Agrawal S,
5. Bandelt HJ,
6. Yao YG,
7. Khan F,
8. Zhu CL,
9. Chaudhuri TK,
10. Zhang YP
(2006) Mol Biol Evol 23:683–690.
Abstract/FREE Full Text
43. ↵
1. Kivisild T,
2. Tolk H-V,
3. Parik J,
4. Wang Y,
5. Papiha SS,
6. Bandelt H-J,
7. Villems R
(2002) Mol Biol Evol 19:1737–1751, (erratum 20:162).
Abstract/FREE Full Text
44. ↵
1. Tanaka M,
2. Cabrera VM,
3. Gonzalez AM,
4. Larruga JM,
5. Takeyasu T,
6. Fuku N,
7. Guo L-J,
8. Hirose R,
9. Fujita Y,
10. Kurata M,
11. et al.
(2004) Genome Res 14:1832–1850.
Abstract/FREE Full Text
45. ↵
1. Macaulay V,
2. Hill C,
3. Achilli A,
4. Rengo C,
5. Clarke D,
6. Meehan W,
7. Blackburn J,
8. Semino O,
9. Scozzari R,
10. Cruciani F,
11. et al.
(2005) Science 308:1034–1036.
Abstract/FREE Full Text
46. ↵
1. Kivisild T,
2. Rootsi S,
3. Metspalu M,
4. Mastana S,
5. Kaldma K,
6. Parik J,
7. Metspalu E,
8. Adojaan M,
9. Tolk H-V,
10. Stepanov V,
11. et al.
(2003) Am J Hum Genet 72:313–332.
CrossRefMedlineISI
47. ↵
1. Hurles ME,
2. Sykes BC,
3. Jobling MA,
4. Forster P
(2005) Am J Hum Genet 76:894–901.
CrossRefMedlineISI
48. ↵
1. Forster P
(2004) Philos Trans R Soc London B 359:255–264.
CrossRefISI
49. ↵
1. Röhl A,
2. Brinkmann B,
3. Forster L,
4. Forster P
(2001) Int J Legal Med 115:29–39.
CrossRefMedlineISI
50. ↵
1. Forster P,
2. Torroni A,
3. Renfrew C,
4. Röhl A
(2001) Mol Biol Evol 18:1864–1881.
Abstract/FREE Full Text
51. ↵
1. Trejaut JA,
2. Kivisild T,
3. Loo JH,
4. Lee CL,
5. He CL,
6. Hsu CJ,
7. Lee ZY,
8. Lin M
(2005) PLoS Biol 3:e247.
CrossRefMedline
52. ↵
1. Stoneking M,
2. Jorde LB,
3. Bhatia K,
4. Wilson AC
(1990) Genetics 124:717–733.
Abstract
53. ↵
1. Evans N,
2. Jones R
1. McConvell P,
2. Evans N
in Archaeology and Linguistics: Aboriginal Australia in Global
Perspective, eds McConvell P, Evans N (Oxford Univ Press, Melbourne,
Australia), pp 385–418.
54. ↵
1. Crystal D
(1987) The Cambridge Encyclopedia of Language (Cambridge Univ
Press, Cambridge, UK).
55. ↵
1. Mellars P
(2006) Science 313:796–800.
Abstract/FREE Full Text
56. ↵
1. Karafet T,
2. Xu L,
3. Du R,
4. Wang W,
5. Feng S,
6. Wells RS,
7. Redd AJ,
8. Zegura SL,
9. Hammer MF
(2001) Am J Hum Genet 69:615–628.
CrossRefMedline
57. ↵
1. Sanchez JJ,
2. Endicott P
(2006) Nat Protocols 1:1370–1378.
CrossRef
58. ↵
1. Endicott P,
2. Metspalu M,
3. Stringer C,
4. Macaulay V,
5. Cooper A,
6. Sanchez JJ
(2006) PLoS ONE 1:e81.
CrossRef
59. ↵
1. Bandelt H-J,
2. Forster P,
3. Sykes BC,
4. Richards MB
(1995) Genetics 141:743–753.
Abstract
60. ↵
1. Bandelt H-J,
2. Forster P,
3. Röhl A
(1999) Mol Biol Evol 16:37–48.
Abstract
61. ↵
1. Forster P,
2. Harding R,
3. Torroni A,
4. Bandelt H-J
(1996) Am J Hum Genet 59:935–945.
MedlineISI
62. ↵
1. Saillard J,
2. Forster P,
3. Lynnerup N,
4. Bandelt H-J,
5. Nørby S
(2000) Am J Hum Genet 67:718–726.
CrossRefMedlineISI
* Add to CiteULikeCiteULike
* Add to ComploreComplore
* Add to ConnoteaConnotea
* Add to Del.icio.usDel.icio.us
* Add to DiggDigg
What's this?
Articles citing this article
*
o Y. Moodley,
o B. Linz,
o Y. Yamaoka,
o H. M. Windsor,
o S. Breurec,
o J.-Y. Wu,
o A. Maady,
o S. Bernhoft,
o J.-M. Thiberge,
o S. Phuanukoonnon,
o G. Jobb,
o P. Siba,
o D. Y. Graham,
o B. J. Marshall,
o and M. Achtman
The Peopling of the Pacific from a Bacterial PerspectiveScience
2009 323:527-530
o Abstract
o Full Text
o Full Text (PDF)
*
o J. S. Lansing,
o J. C. Watkins,
o B. Hallmark,
o M. P. Cox,
o T. M. Karafet,
o H. Sudoyo,
o and M. F. Hammer
Male dominance rarely skews the frequency distribution of Y
chromosome haplotypes in human populationsProc. Natl. Acad. Sci. USA
2008 105:11645-11650
o Abstract
o Full Text
o Full Text (PDF)
*
o P. Soares,
o J. A. Trejaut,
o J.-H. Loo,
o C. Hill,
o M. Mormina,
o C.-L. Lee,
o Y.-M. Chen,
o G. Hudjashov,
o P. Forster,
o V. Macaulay,
o D. Bulbeck,
o S. Oppenheimer,
o M. Lin,
o and M. B. Richards
Climate Change and Postglacial Human Dispersals in Southeast
AsiaMol Biol Evol 2008 25:1209-1218
o Abstract
o Full Text
o Full Text (PDF)
*
o T. M. Karafet,
o F. L. Mendez,
o M. B. Meilerman,
o P. A. Underhill,
o S. L. Zegura,
o and M. F. Hammer
New binary polymorphisms reshape and increase resolution of the
human Y chromosomal haplogroup treeGenome Res 2008 18:830-838
o Abstract
o Full Text
o Full Text (PDF)









.jpg)